Mayor prevalencia de leucemia eritroide aguda en residentes de gran altitud (4000 m)
Higher prevalence of acute erythroid leukemia in high-altitude dwellers (4000 m)
Amaru R1,2; Mamani LF1; Luna J1; Quispe T1; Valencia J C1; Carrasco M2; Patón D1; Mancilla S1; Amaru A4
1. Unidad de Biología Celular, Facultad de Medicina, Universidad Mayor de San Andrés, La Paz, Bolivia
2. Instituto de Oncohematología, La Paz, Bolivia
3. Oncoclinic, Clínica Oncológica, La Paz, Bolivia
Amaru R 0000-0003-2765-9748
Mamani LF 0009-0007-8285-0584
Luna J 0000-0003-3702-157X
Quispe T 0000-0002-9300-2608
Valencia JC 0000-0002-0434-9766
Carrasco M 0009-0006-5399-563X
Patón D 0000-0002-8140-4351
Mancilla S 0009-0009-0869-1305
Amaru A 0000-0003-4170-3266
Palabras clave: leucemia eritroide aguda; eritroleucemia; gran altitud
Keywords: acute erythroid leukemia; erythroleukemia; high altitude
RESUMEN
Introducción. La leucemia eritroide aguda (LEA) es un subtipo raro y agresivo de leucemia mieloide aguda, asociada a mutaciones bialélicas del gen TP53. La edad media de diagnóstico es 67 años conllevando una proporción hombre:mujer de 2,4:1. La hipoxia y los factores inducibles por hipoxia (HIF) parecen desempeñar un rol en el desarrollo de LEA mediante mecanismos que involucran la diferenciación eritroide, producción de hemoglobina, regulación de eritropoyesis y modificación del microambiente de la médula ósea. Por lo que, para discernir sobre la influencia de la hipoxia en este subtipo de leucemia, es de interés analizar su prevalencia y características en poblaciones residentes a gran altitud.
Material y Métodos. Estudio retrospectivo que consideró casos de LMA (n=211) diagnosticados en Bolivia durante el periodo de mayo de 2019 a octubre de 2024, se identificó casos del subtipo LEA (n=15) y se recopiló datos demográficos, hematológicos, morfológicos e inmunofenotípicos. El análisis estadístico contempló categorizar los casos de LMA y subsecuentemente de LEA según la altitud de presentación: 4000m, 2000m y 400m.
Resultados. Los casos de LEA en Bolivia correspondieron al 7,5 % entre todos los casos de LMA, la edad media de presentación fue 53 años con predominio en varones (1,5:1). Distribuidos por altitudes, los casos de LEA a 4000 m (n=10) representaron el 14,1 %, significativamente mayor (p=0,01) comparado con el 4,5 % observado en los casos a 2000 m (n=3) y 2,7 % en los casos a 400 m (n=2). Los índices hematológicos de LEA reflejaron hemoglobina media de 7,3 g/dl, leucocitos 10266/ul y plaquetas 82267/ul, sin diferencias representativas entre las diferentes altitudes. Los rasgos de médula ósea reflejaron >80% de precursores eritroides prominentes con núcleos grandes e irregulares, cromatina dispersa, 1 a 3 nucléolos alargados, citoplasma profundamente basófilo e intensa actividad mitótica. Las características inmunofenotípicas mostraron clonalidad eritroide 90 % (CD34, CD71, CD105, CD36, CD235) y clonalidad mieloide 1,4 % (CD117, CD34).
Conclusiones El porcentaje de LEA (14,1 %), respecto del total de casos de LMA, está aumentada en la altitud (4000 m). Tal aumento podría deberse al incremento de HIF y Eritropoyetina en ambientes de hipoxia hipobárica.
ABSTRACT
Introduction. Acute erythroid leukemia (AEL) is a rare and aggressive subtype of acute myeloid leukemia (2 %) associated with biallelic TP53 mutations. Diagnosis median age stands 67 years old with a 2.4:1 male-to-female proportion. To this, hypoxia and the hypoxia-inducible factors (HIFs) seem to play a role in AEL development through mechanisms involving erythroid differentiation, hemoglobin production, erythropoiesis regulation as well as bone marrow microenvironment modification. Thus, to discern the influence of hypoxia regarding this leukemia subtype, we searched for the prevalence of AEL and its features in the Andean population living at high altitude (4000 m).
Material and Methods. Retrospective study that considered cases of AML (n=211) diagnosed in Bolivia from the period of May 2019 to October 2024, of which AEL cases (n=15) were identified and demographics, hematologic, bone marrow morphology, and immunophenotypic features were gathered. AML cases and subsequently AEL cases were categorized and statistically analyzed according to the altitude of presentation: 4000m, 2000m, and 400m.
Results. AEL in Bolivia corresponded to 7.5 % of all AML cases, median age was 53 years, and predominant in male (1.5:1). At different altitudes, AEL cases at 4000 m (n=10) accounted for 14.1%, significantly higher (p=0.01) when compared to cases at 2000 m (n=3) 4.5 % and cases at 400 m (n=2) 2.7 %. AEL hematological indices did not differ among altitude groups, mean of Hb 7.3 g/dl, WBC: 10266/ul and Plt: 82267/ul. Bone marrow findings reflected >80% of prominent erythroid precursors with large irregular nuclei, dispersed chromatin, 1 to 3 elongated nucleoli, deeply basophilic cytoplasm, and an intense mitotic activity. Immunophenotypic features displayed erythroid clonality 90% (CD34, CD71, CD105, CD36, CD235) and myeloid clonality 1,4% (CD117, CD34).
Conclusions. Among all AML cases, AEL prevalence percentage (14,1) is increased at high altitude (4000 m). Such increase may be due to the increase in HIF and erythropoietin at hypobaric hypoxia environment, so studying the mechanisms involved are of interest.
INTRODUCCIÓN
La leucemia eritroide aguda (LEA), también conocida como leucemia eritroide pura, es un subtipo raro y único de leucemia mieloide aguda (LMA) caracterizada por la proliferación descontrolada de precursores eritroides que conducen a una acumulación de glóbulos rojos inmaduros y anormales en la médula ósea y la sangre periférica(1) con una alta frecuencia de ganancias y amplificaciones que involucran la vía del EPOR/JAK2(2, 3), fosforilación de ERK1/2 al inhibir la actividad del promotor Fli-1 (3, 4) y mutaciones bialélicas TP53(3, 5, 6).
La LEA representa el 2 % de los casos de leucemia mieloide aguda(7-10) y puede evolucionar a partir de un síndrome mielodisplásico previo o desarrollarse de novo. Es un subtipo agresivo de pronóstico malo (supervivencia media global de <6 meses) que se observa por lo habitual en adultos con una edad media de entre 66 y 68 años(8, 11), con un predominio leve en varones (2,4:1)(3). Su distinción se atribuye a la proliferación eritroblástica (>80 %), pancitopenia y afectación extensa de la médula ósea por proeritroblastos (≥30 %). Los marcadores inmunofenotípicos comunes incluyen CD105, CD34, CD71, CD36, CD235(12).
Entre tanto, otros estudios han reportado que la hipoxia influye en el desarrollo de LEA a través de varios mecanismos, incluida la diferenciación eritroide, la producción de hemoglobina, la regulación de la eritropoyesis y la modificación del microambiente de la médula ósea (13, 14). En ello, los reguladores principales que median las respuestas celulares a la hipoxia son los factores inducibles por hipoxia (HIF). Estos complejos heterodímeros están compuestos por subunidades HIF-alfa reguladas por oxígeno (HIF-1alfa, HIF-2alfa) y la subunidad HIF-beta expresada de forma constitutiva (HIF-1 beta)(15). El HIF-1 alfa participa principalmente en la respuesta inicial a la hipoxia aguda, mientras que el HIF-2 alfa responde a la exposición crónica(16, 17). Asimismo, incluso en hipoxia prolongada, el HIF-1 alfa sufre un control de retroalimentación, mientras que el HIF-2 alfa se estabiliza(18). En consecuencia, el aumento de la expresión de HIF se correlaciona con el crecimiento tumoral y la resistencia a las terapias, lo que conduce a la recaída de la enfermedad(19), por lo que, la señalización mediada por HIF constituye un factor relevante en las leucemias(20).
HIF-1 alfa promueve la entrada del G0 al G1 y disminuye la fase S en las células de la LMA a través de la regulación positiva de p27(20, 21); por el contrario, HIF-2 alfa desencadena la progresión y proliferación del ciclo celular al aumentar c-MYC/MAX y su actividad, desempeñando un papel crucial en los procesos leucemogénicos(21, 22). Así, la sobreexpresión de los HIFs está relacionada con un mal pronóstico en LMA,(14) el HIF-1alfa está involucrado en etapas tempranas del cáncer, mientras que el HIF-2 alfa en etapas más tardías(23). Se conoce que HIF1alfa se sobreexpresa en tipos específicos de cáncer humano, incluidas las células leucémicas(24), y que la expresión de HIF-2 alfa juega un papel importante en la regulación de la proliferación en células eritroleucémicas en condiciones de hipoxia(25).
De esta manera, para discernir sobre la influencia de la hipoxia en la leucemia, particularmente de este subtipo tan agresivo de LMA, fue de interés indagar sobre la prevalencia y principales características de la LEA en un ambiente de hipoxia hipobárica, propiamente en la población andina de Bolivia residente a una altitud de 4000 m s.n.m.
MATERIAL Y MÉTODOS
Estudio retrospectivo de corte transversal que consideró analizar casos de LMA (n=211) diagnosticados en Bolivia, durante el período de mayo de 2019 a octubre de 2024, esto considerando las diferentes altitudes de presentación: 4000m (n=71), 2000m (n=67) y 400m (n=73). Los diagnósticos fueron realizados en la Unidad de Biología Celular, centro de referencia nacional para el diagnóstico de enfermedades oncohematológicas. Se corroboró que los diagnósticos de LMA fueron realizados según el protocolo de Euroflow(26).
Consecutivamente, con base en los criterios de la Organización Mundial de la Salud del 2022(27), se identificó casos del subtipo LEA. Se recopiló reportes de las características demográficas, de altitud, hematológicas, morfológicas e inmunofenotípicas.
Las placas del estudio citomorfológico (frotis de médula ósea) y datos de citometría de flujo disponibles para su revisión a fin de confirmar el diagnóstico de LEA constituyeron los criterios de inclusión de casos. Se corroboró que los frotis de médula ósea fueron realizados siguiendo protocolos de tinción con May-Grunwald-Giemsa y la inmunofenotipificación fue realizada con citómetro de flujo BD Canto II empleando anticuerpos CD45, CD34, CD117, CD71, CD105, CD36, CD235.
Este estudio estuvo adherido a los principios establecidos en la Declaración de Helsinki y obtuvo aprobación del Comité de Revisión de la Facultad de Medicina de la Universidad Mayor de San Andrés.
Los datos fueron sistematizados utilizando Excel 16.29.1. y subsecuente análisis descriptivo, la proporción de LEA en diferentes altitudes se obtuvo mediante Chi cuadrado, se consideró un valor p de significancia <0.05.
RESULTADOS
Entre los 211 diagnósticos de LMA, se identificó 15 casos de LEA (Tabla 1) que correspondieron al 7,5 %, la edad media de presentación fue de 52,6 años con un ligero predominio en varones (n=9) evidenciándose una proporción hombre-mujer de 1,5:1.
En cuanto a las características inmunofenotípicas, se evidenció 90 % de clonalidad eritroide (CD34, CD71, CD105, CD36, CD235) y 1,4 % de clonalidad mieloide (CD117, CD34), Tabla 1. Los rasgos percibidos en la médula ósea reflejaron >80 % de precursores eritroides prominentes con núcleos grandes e irregulares, cromatina dispersa, 1 a 3 nucléolos alargados, citoplasma profundamente basófilo y una intensa actividad mitótica (Figura 1).
Tabla 1. Características inmunofenotípicas y hematológicas de LEA en gran altitud
|
Sexo |
Edad años |
Altitud msnm |
Clonalidad eritroide Médula ósea |
Clonalidad mieloide Médula ósea |
Hb g/dl |
GB /ul |
PLT 103/ul |
|
F |
32 |
400 |
83% CD34/71/105 |
2% CD34/11 |
6.3 |
50000 |
70 |
|
F |
71 |
400 |
88% CD34/71/105/36/235 |
1% CD34/117 |
6.8 |
4500 |
67 |
|
F |
39 |
2000 |
94% CD34/71/105/36 |
3% CD34/117 |
6.2 |
1830 |
81 |
|
M |
72 |
2000 |
89% CD71/105/36/235 |
1% CD34/117 |
5.4 |
5300 |
66 |
|
M |
48 |
2000 |
89% CD71/105/36/235 |
1% CD34/117 |
6.2 |
2000 |
144 |
|
M |
56 |
4000 |
82% CD34/71/105/235 |
1% CD34/117 |
--- |
3300 |
60 |
|
F |
73 |
4000 |
87% CD34/71/36/235 |
1% CD34/117 |
10 |
1500 |
150 |
|
M |
65 |
4000 |
97% CD34/71/36/235 |
2% CD34/117 |
7.9 |
58300 |
84 |
|
M |
50 |
4000 |
98% CD34/71/105/36/235 |
1% CD34/117 |
8.3 |
3700 |
170 |
|
F |
55 |
4000 |
92% CD34/71/105/36/235 |
1% CD34/117 |
7.2 |
1900 |
48 |
|
M |
26 |
4000 |
97% CD34/71/105/36/235 |
1% CD34/117 |
7.9 |
2000 |
49 |
|
M |
58 |
4000 |
89% CD71/105/36/235 |
1% CD34/117 |
8.0 |
11260 |
74 |
|
M |
37 |
4000 |
95% CD71/105/36/235 |
2% CD34/117 |
7.2 |
2500 |
54 |
|
M |
64 |
4000 |
90% CD71/105/235 |
1% CD34/117 |
7.1 |
3600 |
50 |
|
F |
43 |
4000 |
80% CD71/105/36/235 |
1% CD34/117 |
9.8 |
2300 |
67 |
Figura 1. Características morfológicas de LEA
|
Blastos eritroides |
Actividad mitótica |
|
|
|
|
|
|
|
|
|
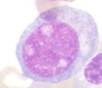
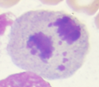
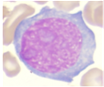
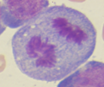
Así también, al analizar los casos de LEA distribuidos por altitudes (Tabla 2), se evidenció 10 casos a 4000 m (3 mujeres, 7 hombres, edad media de 53 años) que representaron el 14,1%, el cual fue significativamente mayor (p = 0,01) comparado con 3 casos a 2000 m (2 hombres, 1 mujer, edad media de 53 años) correspondiendo al 4,5 % y 2 casos a 400 m (mujeres, edad media de 52 años) que representaron el 2,7 %.
Tabla 2. Proporción de Leucemia Eritroide Aguda en diferentes altitudes
|
Ciudad y altitud |
LMA total (n=) |
LEA (n=) |
LEA (%) |
p= |
|
Santa Cruz, 400 msnm |
73 |
2 |
2.7 |
0.018* |
|
Cochabamba, 2000 msnm |
67 |
3 |
4.5 |
|
|
La Paz, 4000 msnm |
71 |
10 |
14.1 |
* Valor de chi cuadrado 8.04 con 2 grados de libertad, valor p= 0.018
Los índices hematológicos del total de casos de LEA mostraron una media de Hb de 7,3 g/dl, leucocitos 10266/ul y plaquetas 82267/ul. De la misma manera, considerando los casos en la altitud (4000 m), la media de Hb fue de 7,9 g/dl, leucocitos 9036/ul y plaquetas 80600/ul, estos valores no difirieron con los promedios de las otras altitudes (2000 m, 400 m).
DISCUSIÓN
Conforme los resultados obtenidos en este estudio, se reporta que la proporción de LEA en la población andina residente a una altitud de 4000 m correspondió al 14,1 % del total de casos de LMA, porcentaje significativamente mayor (p=0,01) al ser comparado con los casos en otras altitudes (4,5 % a 2000 m y 2,7 % a 400 m). Este porcentaje a gran altitud difiere significativamente del 2 % reportado en otros estudios a nivel mundial; asimismo, la media de edad del diagnóstico (53 años) refleja ser menor contrastado con otros reportes (>65 años)(3, 7-10). Sin embargo, el predominio en varones (1,5:1) es similar a lo reportado en otros estudios(3). Por su parte, las características morfológicas e inmunofenotípicas de LEA observadas en nuestro estudio concuerdan con las características convencionales(28).
Los seres humanos que habitan grandes altitudes en condiciones de hipóxia hipobárica tienen un HIF aumentado(29); y como consecuencia de la estimulación del HIF, la concentración plasmática de eritropoyetina (EPO) (30) y la expresión del JAK2 aumentan rápidamente en esas condiciones(31). Referente al eje HIF-EPO-JAK2, recientemente se ha reportado mutaciones de los genes EPO y JAK2 asociados a AEL(32). En la leucemia mieloide aguda (LMA), los niveles de proteínas HIF-1 alfa y HIF-2 alfa están aumentados al compararlos con los leucocitos normales de la médula ósea, y por su parte, se encontró que la expresión de HIF-2 alfa está aumentada en LEA (33, 34). También, se ha reportado que en las células de LMA, que portan mutaciones TP53, los genes diana HIF-1alfa están regulados positivamente(34, 35) donde también una actividad anormal de la proteína de unión GATA1 está implicada(34, 35).
Se considera que la actividad alterada de GATA1 desempeña un papel importante en la biología molecular de LEA a través de los diversos efectos sobre la línea eritroide y la inhibición de la diferenciación eritroide normal(3). La inducción de GATA1 humana mediada por HIF-1alfa promueve la diferenciación eritroide en condiciones hipóxicas(36); y, en presencia de TP53 mutado, caracterizado por una pérdida de función bialélica de TP53, coopera con múltiples vías alternativas para producir el fenotipo LEA(37). Por lo tanto, se puede suponer que un entorno hipóxico hipobárico induce un aumento de HIF-alfa y modifica GATA1 activando una proliferación eritroide anormal en los pacientes con pérdida de la función de P53.
De este modo, alteraciones en las actividades del eje HIF-EPO-JAK2 y GATA 1 en condiciones de hipoxia hipobárica estarían relacionadas con una eritropoyesis alterada y, en combinación con la mutación bialélica del P53, subsecuentemente desencadenar el desarrollo de LEA. Esta posible interacción molecular podría correlacionarse con la proporción aumentada del subtipo LEA en la población residente en gran altitud, según reflejan los resultados obtenidos en este estudio. De manera que, estudios posteriores sobre los mecanismos moleculares involucrados en dicha interacción requieren ser confirmados, conjuntamente a estudios con mayor número de casos y considerando más variables epidemiológicas.
LIMITACIONES DEL ESTUDIO
Tamaño muestral pequeño y datos clínicos insuficientes para analizar otros factores o variables epidemiológicas como trastornos hematológicos previos, exposición a tóxicos, etc.
Contribución de los autores: todas las personas autoras han efectuado una contribución sustancial a la concepción o el diseño del estudio o a la recolección, análisis o interpretación de los datos; han participado en la redacción del artículo o en la revisión crítica de su contenido intelectual; han aprobado la versión final del manuscrito; y son capaces de responder respecto de todos los aspectos del manuscrito de cara a asegurar que las cuestiones relacionadas con la veracidad o integridad de todos sus contenidos han sido adecuadamente investigadas y resueltas.
Referencias
1. Zhang W, Yamamoto K, Chang Y-H, y col. HDAC7 is a potential therapeutic target in acute erythroid leukemia. Leukemia. 2024:1-14.
2. Takeda J, Yoshida K, Nakagawa MM, y col. Amplified EPOR/JAK2 Genes Define a Unique Subtype of Acute Erythroid Leukemia. Blood Cancer Discov. 2022;3(5):410-27 doi: 10.1158/2643-3230.Bcd-21-0192.
3. Fernandes P, Waldron N, Chatzilygeroudi T, Naji NS, Karantanos T. Acute Erythroid Leukemia: From Molecular Biology to Clinical Outcomes. International Journal of Molecular Sciences. 2024;25(11):6256.
4. Mo M, Ma Y, Yu J, y col. A Novel Chalcone Derivative Regulates the Expression and Phosphorylation of ERK1/2 by Inhibiting Fli-1 Promoter Activity for Preventing the Malignant Progression of Erythroleukemia. Catalysts. 2022;13(1):84.
5. Piqué-Borràs M-R, Jevtic Z, Bagger FO, y col. The NFIA-ETO2 fusion blocks erythroid maturation and induces pure erythroid leukemia in cooperation with mutant TP53. Blood. 2023;141(18):2245-60 doi: 10.1182/blood.2022017273.
6. Alexander C. A History and Current Understanding of Acute Erythroid Leukemia. Clinical Lymphoma Myeloma and Leukemia. 2023;23(8):583-8.
7. Sato S, Kobayashi M, Suzaki K, Nanke I, Kosugi N. Challenging Diagnosis of Pure Erythroid Leukemia: A Case Report and Literature Review. Hematology Reports. 2022;14(1):38-44.
8. Reichard KK, Tefferi A, Abdelmagid M, y col. Pure (acute) erythroid leukemia: morphology, immunophenotype, cytogenetics, mutations, treatment details, and survival data among 41 Mayo Clinic cases. Blood Cancer Journal. 2022;12(11):147 doi: 10.1038/s41408-022-00746-x.
9. Ouahidi I, Yahyaoui H, Nassiri Z, Aitameur M, Chakour M. Pure erythroid leukemia De Novo in 16-years-old girl: A case report and literature review. World Journal of Advanced Research and Reviews. 2022;16(3):198-204.
10. Zhang Y, Li K, Li X, Wang H, Li T, Long F. Concurrent Pleural and Pericardial Involvement in a Patient With De Novo Pure Erythroid Leukemia. Ann Lab Med. 2023 doi: 10.3343/alm.2023.0252.
11. Reinig EF, Greipp PT, Chiu A, Howard MT, Reichard KK. De novo pure erythroid leukemia: refining the clinicopathologic and cytogenetic characteristics of a rare entity. Mod Pathol. 2018;31(5):705-17 doi: 10.1038/modpathol.2017.175.
12. Wang W, Wang SA, Medeiros LJ, Khoury JD. Pure erythroid leukemia. American journal of hematology. 2017;92(3):292-6.
13. Bapat A, Schippel N, Shi X, y col. Hypoxia promotes erythroid differentiation through the development of progenitors and proerythroblasts. Exp Hematol. 2021;97:32-46.e35 doi: 10.1016/j.exphem.2021.02.012.
14. Giuntoli S, Rovida E, Gozzini A, y col. Severe hypoxia defines heterogeneity and selects highly immature progenitors within clonal erythroleukemia cells. Stem Cells. 2007;25(5):1119-25 doi: 10.1634/stemcells.2006-0637.
15. Semenza GL. Hypoxia-inducible factors: mediators of cancer progression and targets for cancer therapy. Trends in pharmacological sciences. 2012;33(4):207-14.
16. Koh MY, Powis G. Passing the baton: the HIF switch. Trends in biochemical sciences. 2012;37(9):364-72.
17. Holmquist-Mengelbier L, Fredlund E, Löfstedt T, y col. Recruitment of HIF-1α and HIF-2α to common target genes is differentially regulated in neuroblastoma: HIF-2α promotes an aggressive phenotype. Cancer cell. 2006;10(5):413-23.
18. Concetti J, Wilson CL. NFKB1 and cancer: friend or foe? Cells. 2018;7(9):133.
19. Mimeault M, Batra SK. Hypoxia‐inducing factors as master regulators of stemness properties and altered metabolism of cancer‐and metastasis‐initiating cells. Journal of cellular and molecular medicine. 2013;17(1):30-54.
20. Deynoux M, Sunter N, Hérault O, Mazurier F. Hypoxia and Hypoxia-Inducible Factors in Leukemias. Frontiers in Oncology. 2016;6 doi: 10.3389/fonc.2016.00041.
21. Drolle H, Wagner M, Vasold J, y col. Hypoxia regulates proliferation of acute myeloid leukemia and sensitivity against chemotherapy. Leukemia research. 2015;39(7):779-85.
22. Gordan JD, Bertout JA, Hu C-J, Diehl JA, Simon MC. HIF-2α promotes hypoxic cell proliferation by enhancing c-myc transcriptional activity. Cancer cell. 2007;11(4):335-47.
23. Murugesan T, Rajajeyabalachandran G, Kumar S, Nagaraju S, Jegatheesan SK. Targeting HIF-2α as therapy for advanced cancers. Drug Discov Today. 2018;23(7):1444-51 doi: 10.1016/j.drudis.2018.05.003.
24. Itoh M, Okuhashi Y, Takahashi Y, y col. Hypoxia Up-regulates HIF expression while suppressing cell growth and NOTCH activity in leukaemia cells. Anticancer Research. 2019;39(8):4165-70.
25. Xu QQ, Xiao FJ, Sun HY, y col. Ptpmt1 induced by HIF-2α regulates the proliferation and glucose metabolism in erythroleukemia cells. Biochem Biophys Res Commun. 2016;471(4):459-65 doi: 10.1016/j.bbrc.2016.02.053.
26. Kalina T, Flores-Montero J, Van Der Velden V, y col. EuroFlow standardization of flow cytometer instrument settings and immunophenotyping protocols. Leukemia. 2012;26(9):1986-2010.
27. Alexander C. A History and Current Understanding of Acute Erythroid Leukemia. Clinical Lymphoma Myeloma and Leukemia. 2023.
28. Zuo Z, Polski JM, Kasyan A, Medeiros LJ. Acute erythroid leukemia. Archives of pathology & laboratory medicine. 2010;134(9):1261-70.
29. Pham K, Parikh K, Heinrich EC. Hypoxia and inflammation: insights from high-altitude physiology. Frontiers in physiology. 2021;12:676782.
30. Schuler B, Lundby C, Gassmann M. HIF-1 and the adaptation of man to high altitude. SCHWEIZERISCHE ZEITSCHRIFT FUR SPORTMEDIZIN UND SPORTTRAUMATOLOGIE. 2005;53(2):82.
31. Siques P, Pena E, Brito J, El Alam S. Oxidative Stress, Kinase Activation, and Inflammatory Pathways Involved in Effects on Smooth Muscle Cells During Pulmonary Artery Hypertension Under Hypobaric Hypoxia Exposure. Frontiers in Physiology. 2021;Volume 12 - 2021 doi: 10.3389/fphys.2021.690341.
32. Jena D, Balasubramanian P, Hajra S, Raghav SK. Novel Mutations in Acute Erythroid Leukemia–A Case Report with Review of Literature. Indian Journal of Hematology and Blood Transfusion. 2025;41(2):383-7 doi: 10.1007/s12288-024-01826-7.
33. Forristal C, Brown A, Helwani F, y col. Hypoxia inducible factor (HIF)-2α accelerates disease progression in mouse models of leukemia and lymphoma but is not a poor prognosis factor in human AML. Leukemia. 2015;29(10):2075-85.
34. Magliulo D, Bernardi R. Hypoxic stress and hypoxia-inducible factors in leukemias. Frontiers in Oncology. 2022;12:973978.
35. Wang Y, Liu Y, Bailey C, y col. Therapeutic targeting of TP53-mutated acute myeloid leukemia by inhibiting HIF-1α with echinomycin. Oncogene. 2020;39(14):3015-27.
36. Liu F, Hu C, Ding J, Fu C, Wang S, Li T. GATA-1 Promotes Erythroid Differentiation Through the Upregulation of miR-451a and miR-210-3p Expressions in CD34+ Cells in High-Altitude Polycythemia. High Altitude Medicine & Biology. 2023;24(1):59-67.
37. Fang H, Wang SA, Khoury JD, y col. Pure erythroid leukemia is characterized by biallelic TP53 inactivation and abnormal p53 expression patterns in de novo and secondary cases. Haematologica. 2022;107(9):2232.